
Centre for Human Genetics
Bangalore.
Bangalore.

Prof. Vidyanand Nanjundiah
Ph.D University of Chicago,USA
My broad interest is in understanding the evolution of cooperative behaviour. I have been studying soilamoebae known as the cellular slime moulds. They have a life cycle that has distinct unicellular and multicellular phases. Multicellularity involves cooperative behaviour with division of labour, and is initiated after cells stop dividing once the bacterial food supply runs out. The multicellular structure is like a small society of cells, some of which differentiate into spores that can resist starvation and the rest die, form a vertical stalk and support the spore mass, apparently to aid its passive dispersalto a location where food is present. Thus, stalk cells appear to behave "altruistically", i.e. in a manner that seems to benefit other cells, i.e. the spores,rather than themselves. Experiments carried out to understand this puzzle have led to the following findings. (i) Naturally occurring social groups are genetically heterogeneous; therefore everyone's reproductive interest is not the same. (ii) Within agroup, some amoebae are better at one fitness related task, while other amoebae are better at a different task. (iii) In some cases, the fitness of an amoeba belonging to a certain genotype can be high or low, depending on whether the genotypeis rare or common, respectively (see figure), withinthe group.Therefore, once heterogeneity between groups is taken into account, the overall reproductive advantage of cells belonging to different genotypes can be the same. This is a testable hypothesis for explaining the apparent altruism of stalk cells.
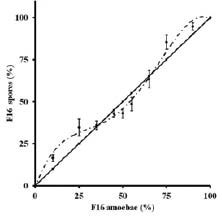
Starved amoebae of two genotypes, F5 and F16 (proportion of F16 on X-axis), were mixed and allowed to form multicellular groups. The proportion of survivors (spores) belonging to F16 is shown on the Y-axis. F16 does better that F5 at certain mixing ratios and worse at others. A stable equilibrium is possible when about 40% of the amoebae belong to F16 (from Sathe and VN, 2018).
Selected publications:
1.Complex interactions underpin social behaviour in Dictyostelium giganteum.Behavioural Ecology and Sociobiology, 72: article number 167 (https://doi.org/10.1007/s00265-018-2572-9) (2018). Sathe S, Nanjundiah V.
2.An individual-level selection model for the apparent altruism exhibited by cellular slime moulds. Journal of Biosciences, 43: 49-58(2018).Zahavi A, Harris KD, Nanjundiah V.
3.Protists and multiple routes to the evolution of multicellularity.In 'History of Evolutionary Cell Biology'. Eds. B Hall and S Moody. CRC Press,Boca Raton (2017). Nanjundiah V, Ruiz-Trillo I,Kirk D.
4.Haldane's view of natural selection.Journal of Genetics 96: 765-772 (2017). Rao V, Nanjundiah V.
5.Cellular slime mold development as a paradigm for the transition from unicellular to multicellular life.In 'Multicellularity: Origins and Evolution'. Eds. KJ Niklas and SA Newman.MIT Press, Cambridge (USA) (2016).Nanjundiah V.
6.Science and Religion. Newsletter, Institute of Bioinformatics and Applied Biotechnology (https://www.ibab.ac.in/wp-content/uploads/ 2019/05/newzine_January_Final-1.pdf) (2019). Nanjundiah V.
7.Does resource availability help determine the evolutionary route to multicellularity?Evolution and Development, 21: 115-119 (2019). Hamant O, Bhat R, Nanjundiah V, Newman SA.
8.John Tyler Bonner (1920-2019).Current Science, 116: 1258-1261 (2019). Nanjundiah V.
9.A response to Lisa Sideris.(to appearin) Christian Theology and Climate Change.Eds. EM Conradie and HP Koster.(2019). Nanjundiah V.
Contact Info
Location
Centre for Human Genetics
Biotech Park, Electronic City Phase 1
Bangalore 560 100
Tel: 91-80-28521832/28521833/28523596
Email: info@chg.res.in
For M.Sc. programme: mscapp@chg.res.in
Copyright © 2011 - All Rights Reserved - Taurus Hard Soft Solutions Pvt. Ltd

Stay Up to Date With Whats Happening